
Shade Preference in Sub-tidal populations of Dorid Nudibranchs: an Analytical Rebuttal
Ryan M. J. Murphy
November, 2000
A Submission for partial fulfillment of the requirements of the International Baccalaureate
PURPOSE:
The purpose of this investigation was to evaluate the claims of H.L. Geiger and A.R. Holyoak (1996) that adult dorid nudibranchs displayed a distinct shade preference during laboratory test runs, and then to analyse their conclusions.
INTRODUCTION:
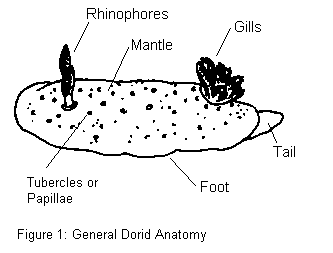
Dorid nudibranchs, oval or ellipsoid in general shape, are dorsoventrally flattened. Protruding from their backs are two sets of structures. Anteriorly is a pair of rhinophores, chemosensory antennae that often have numerous folds and lamellae (or ridges) that serve to increase the sensitive surface area. This is their primary sense organ. More posteriorly are situated the flowerlike, branchial plumes (or gills). Dorids differ from the other main division of nudibranchs, the Aeolids, in that they have no ceratan extensions[1], but rather small papillae or no skin protuberances at all.
(from Morrow, 2000)
Nudibranchs are useful to marine ecologists as they are indicator species, that is, they can be used as early indicators of environmental change, such as degradation, to a community or ecosystem (Pidwirny, 2000). Nudibranchs and other opisthobranchs are useful to neurologists as well. Anisodoris nobilis in particular has been utilised for neurophysiological experiments because of its neurons that are many times larger than human neurons (Smallwood and Rogers, 1908). Also, since nudibranchs have relatively few neurons, neurologists can use nudibranchs to analyse the process by which neuromodulation controls bodily functions to a greater extent than is possible in any vertebrate circuit (Katz, 2000).
Nudibranchs are opisthobranchs, meaning they belong to a group of marine gastropod molluscs of the order Opisthobranchia, characterized by gills, a shell that is reduced or absent, and two pairs of tentacles. Opisthobranchs are simultaneous hermaphrodites, meaning that a single individual has both the male and female gonads (testis and ovary) to make sperm and eggs, as well as the external appendages for the transmission of these sex products by copulation. The final products in copulation of nudibranchs are resilient egg ribbons (Hurst, 1967) with large surface area-to-volume ratios, which are deposited during all seasons of the year. Egg ribbons of the Archidoris montereyensis nudibranch were studied by Biermann et al. (1992), who analysed the effects of solar radiation and other environmental factors on egg deposition site and embryo survival. Biermann et al. (1992) found that A. montereyensis preferentially laid their eggs underneath macroalgal shelter. In its natural habitat and in controlled laboratory conditions, they went on to determine that solar radiation arrested embryo development in tests that exposed the egg ribbons to direct sunlight in a shallow, water-cooled trough. Solar radiation was found to directly or indirectly limit the distribution of nudibranchs and their development in shallow water (Biermann et al., 1992). Biermann et al. proposed that adult dorids may actively select shaded areas to deposit their egg ribbons, an idea forming the basis for a more recent study of dorid nudibranch behaviour (Geiger and Holyoak, 1996).
eiger and Holyoak hypothesised that A. montereyensis adults’ preference to lay egg ribbons in shaded conditions was a direct result of natural selection against offspring of individuals that deposit egg ribbons in unshaded areas (1996). In addition, it was hypothesised that the differential survival of embryos as demonstrated by Biermann et al. (1992) might have an evolutionary effect on reproductive ecology (Geiger and Holyoak, 1996). Time trials conducted in an artificially shaded tank during the late spring involving 3 to 16 specimens of the same species showed that between 83.3 and 100% of the nudibranchs studied preferred shaded conditions to light (Geiger and Holyoak, 1996). Geiger and Holyoak used A. montereyensis specimens from piers at Friday Harbour on San Juan Island (see Friday Harbour, Figure 2), similar to the population studied by Biermann et al. (1992) at Argyle Creek on San Juan Island.
Figure 2: San Juan Island
Figure 3: Pedder Bay-Race Rocks Region
The initial purpose of this investigation was to determine if the shade preference as elucidated by Geiger and Holyoak in A. montereyensis, D. sandiegensis, and T. catalinae was present in other dorid nudibranch species. Five species of nudibranch were chosen for this experiment (see Appendix A): Archidoris montereyensis (MacFarland, 1966), Anisodoris nobilis (MacFarland, 1905), Diaulula sandiegensis (Bergh, 1879), Acanthodoris hudsoni (MacFarland, 1905)and Cadlina luteomarginata (MacFarland, 1905). A. montereyensis and D. sandiegensis were used as a comparison to the Geiger and Holyoak experiment. Cadlina luteomarginata, Acanthodoris hudsoni,and Diaulula sandiegensis were chosen for their local abundance, as well as to determine whether there are behavioural differences between dorids generally more common in the low inter-tidal ranges (i.e. A. hudsoni, A. montereyensis, and D. sandiegensis) and those more common in the subtidal range (i.e. C. luteomarginata and A. nobilis) (Morris et al., 1983). Upon initial tests with A. montereyensis, it became clear that shade preference of other dorids could not be the focus of the investigation, for as Geiger and Holyoak found a negative relationship between dorid movement and light intensity, my preliminary work showed the contrary. The obvious question then arose: Why the population of A. montereyensis and D. sandiegensis specimens being tested did not express shade preference as the Friday Harbour populations did. Subsequent analyses were designed to verify these results, and to help explain this difference.
All five nudibranch species used in this study were collected at Great Race Rocks, 48o17’45” N, 123 o31’50” W (see Figure 3) at depths between 6 and 12 m, subtidal range. Initial time trials were conducted in mid-May, 2000, and all secondary trials were conducted in late October and early November 2000.